Esquema Z
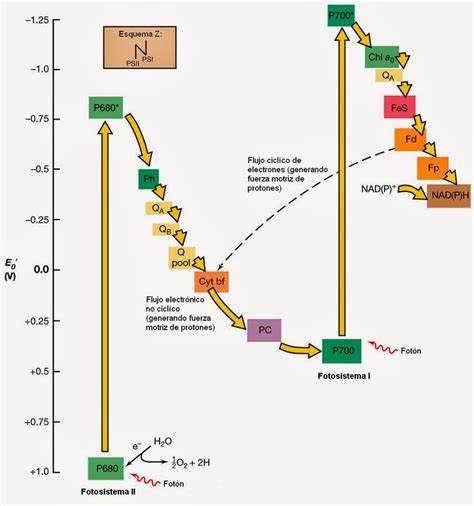
La fase luminosa, fase clara, fase fotoquímica o reacción de Hill es la primera fase de la fotosíntesis, que depende directamente de la luz o energía lumínica para poder obtener energía química en forma de ATP y NADPH, a partir de la disociación de moléculas de agua, formando oxígeno e hidrógeno. La energía creada en esta fase, será utilizada durante la fase oscura, para de esta forma continuar con la fotosíntesis.
Este proceso se realiza en la cadena transportadora del cloroplasto, en los complejos clorofila-proteína que se agrupan en unidades llamadas fotosistemas que están en los tilacoides (membranas internas) de los cloroplastos.
Existen dos tipos de fotosistemas y funcionan gracias a los pigmentos que son los que se encargan de captar la luz, como las clorofilas a y b, o los carotenoides, los cuales absorben diferentes longitudes de onda, formando así tanto al fotosistema I, como al fotosistema II.
En organismos eucariontes fotosintéticos, la fotosíntesis ocurre en el orgánulo subcelular conocido como cloroplasto. El cloroplasto está compuesto por un extenso sistema de membranas internas llamados tilacoides. Los tilacoides contienen clorofila y es en estos donde tienen lugar las reacciones de la fase luminosa. Las reacciones de reducción de carbono o fase oscura tienen lugar en el estroma. La mayoría de los tilacoides están estrechamente asociados entre sí. Cuando se apilan se llaman grana y cuando no se llaman lamelas del estroma.
Los cloroplastos suelen estar rodeados por dos membranas separadas que se conocen como la envoltura. Este sistema de doble membrana contiene diversos sistemas de transporte de metabolitos. Además los cloroplastos contienen su propio DNA, RNA y ribosomas por lo que la mayoría de las proteínas del cloroplasto son producto de la traducción y transcripción del propio cloroplasto, aunque algunas sí son codificadas por el DNA nuclear.

Embebidas en la membrana de los tilacoides se encuentran una gran variedad de proteínas vitales para la fotosíntesis, estas suelen encontrarse en las regiones acuosas a ambos lados del tilacoide. Estas proteínas integrales de membrana están formadas, mayormente, por aminoácidos hidrofóbicos. Los centros de reacción, los complejos proteína-pigmento de las antenas y la mayoría de las enzimas de transporte electrónico son proteínas integrales de membrana.
Tanto el centro de reacción del fotosistema II, como sus clorofilas y proteínas asociadas se encuentran mayormente en los grana. En cambio el centro de reacción del fotosistema I, sus pigmentos antena asociados, las proteínas de transferencia de electrones y la enzima que cataliza la formación de ATP se encuentran casi exclusivamente en las lamelas del estroma y los extremos de los grana.
Esto significa que los dos eventos fotoquímicos implicados en la fotosíntesis aeróbica están muy separados (muchas decenas de nanómetros). Esto nos indica que existen transportadores de electrones entre la membrana en la región del grana a la membrana en la región del estroma. No está clara la razón funcional de esta separación, aunque se cree que es una manera de mejorar la eficiencia de distribución energética entre los dos fotosistemas.
Conforman a la antena colectora de luz, que forma parte junto con el centro de reacción química a un fotosistema. Este complejo está formado por moléculas de pigmento clorofilas a y b, que están ligados a proteínas y algunos carotenoides. Por ejemplo el complejo captador de luz que se encuentra en el fotosistema ll llamado LHC-ll, está formado por:
La luz recolectada por dicho complejo está formada por fotones, la energía de un fotón, absorbida en cualquier punto del conjunto de moléculas de clorofila de la antena, migra a un centro de reacción que promueve el evento de transferencia de un electrón. La energía de los fotones absorbida por los pigmentos (clorofilas)”antena” es transferida por resonancia inductiva (como el de los enlaces covalentes) hasta los centros de reacción. Al haber muchos pigmentos en la antena hacen que se absorba luz con diferente longitud de onda lo que permite un mayor espectro de absorción por lo que la fotosíntesis se hace más eficaz . Por ejemplo la clorofila P680 en el fotosistema II y clorofila P700 en el fotosistema I.
Para que se realice la fotosíntesis es necesario que la energía de los electrones excitados de varios pigmentos se transfiera a un pigmento colector de energía, el cual se denomina Centro de reacción. Un centro de reacción está formado por el pigmento diana, el primer aceptor de electrones y el primer dador de electrones. En la fotosíntesis hay dos centros de reacción el del fotosistema I y el del fotosistema II. Se dice que la energía migra de un pigmento a otro hasta llegar al centro de reacción. Estos centros de reacción transfieren un electrón “rico” en energía que llegó al pigmento vale receptor Ejemplo: Feofitina en el PSII y A0 en el PSI, respectivamente; y reciben un electrón “pobre” en energía del donador Ejemplo: residuo de tirosina en el PSII y plastocianina en el PSI, respectivamente.
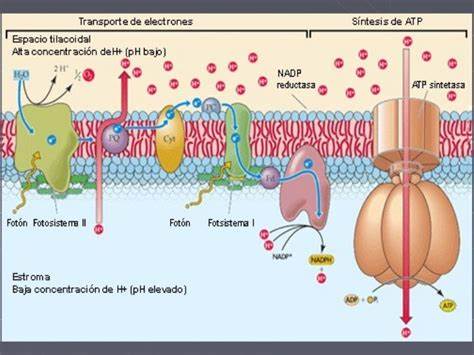
El fotosistema I (PSI) y el fotosistema II (PSII) son los encargados de captar la luz y de emplear su energía para impulsar el transporte de electrones a través de una cadena de aceptores. El complejo antena de dichos fotosistemas atrapa fotones de la luz, elevando los electrones a niveles más altos que su estado cuántico fundamental, y esta energía se va transportando entre diferentes moléculas de clorofila por resonancia, hasta que en el centro del fotosistema II se produce la fotólisis del agua, rompiéndola en medio, originando O, 2 protones y dos electrones. El oxígeno se unirá con el sobrante de otra molécula de agua, para crear oxígeno atmosférico (O
2); los protones translocados al interior del tilacoide contribuyen a crear un gradiente electroquímico, que será utilizado por la ATPasa, y los electrones repondrán la carencia electrónica de la clorofila original.
Para que se pueda absorber la energía lumínica, se requiere de la excitación de los electrones en dos diferentes sitios: los fotosistemas I y II.
Los fotosistemas son el conjunto de proteínas transmembranales que engloban a los pigmentos fotosintéticos cuya función es captar la energía lumínica para utilizarla durante el transporte de electrones y protones.
La distribución de los fotosistemas I y II, así como de la ATP sintasa y el citocromo b/f, no es aleatoria:
Esta separación de los fotosistemas previene que la energía absorbida por el fotosistema II sea transferida directamente el fotosistema I además de posicionar al fotosistema I en regiones donde es más accesible el NADP+, presente en el estroma.
Los eventos de excitación en el fotositema I ocurren de igual manera que en el fotosistema II:

Existen otros dos complejos de proteínas que no están unidos a clorofilas en las membranas de los tilacoides: el complejo del citocromo b6f y el complejo de la ATP-sintasa.
El citocromo b6f es un intermediario en el transporte de electrones entre el fotosistema II y el fotosistema I, capaz de crear un gradiente de energía química que será empleado por el complejo ATP sintasa para generar ATP, en un proceso llamado fotofosforilación. La función del fotosistema II está asociada a la descomposición (fotolisis) de las moléculas de agua (H
2O) en 2 protones (H+
) y O
2. Los dos electrones que procedían de los átomos de hidrógeno de la molécula de agua son captados por el llamado centro de reacción del fotosistema II (P680), elevados a un nivel energético superior por la energía que proporciona la luz, captada por el fotosistema II y una serie de proteínas asociadas a clorofilas y otros pigmentos fotosintéticos (carotenoides) llamadas complejos antena. Desde el nivel energético más alto el electrón puede ir "descendiendo" (como el agua almacenada en una presa) hacia estados energéticos más bajos a través de una cadena transportadora de electrones en la que participan una molécula denominada plastoquinona, el complejo del citocromo b6f y una proteína denominada plastocianina.
Durante este camino parte de la energía del electrón es destinada (por mediación del citocromo b6f) a crear un gradiente de energía a través de las membranas de los tilacoides que será empleado para la síntesis de ATP por la ATP sintetasa.
El proceso nuevo, tiene analogía con la producción de electricidad durante el paso del agua almacenada en una presa a través de las turbinas. La plastocianina transporta los electrones hasta el fotosistema I, que también posee un centro de reacción (P700) y un complejo antena asociado para la captación de luz.
Los electrones que llegan a PSI son de nuevo impulsados por la energía de la luz a un nivel energético superior y también transportados a través de una nueva cadena de aceptores hasta llegar a una molécula final aceptora, el NADP+. Esta molécula, que capta finalmente los electrones será empleada, junto al ATP producido, en la fase posterior de la fotosíntesis ciclo de Calvin para convertir el dióxido de carbono atmosférico (o disuelto en el agua en medios acuáticos) en materia orgánica.
Escribe un comentario o lo que quieras sobre Esquema Z (directo, no tienes que registrarte)
Comentarios
(de más nuevos a más antiguos)
Aún no hay comentarios, ¡deja el primero!