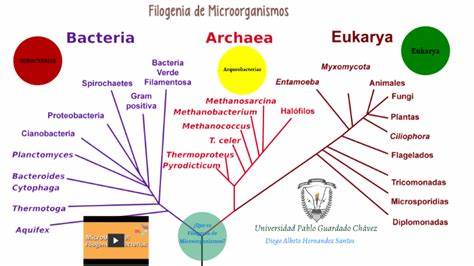
Filogenia bacteriana
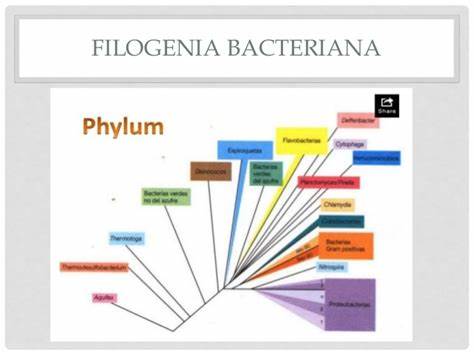
La filogenia de las bacterias se desarrolla en la actualidad a partir de la elaboración de árboles filogenéticos moleculares, especialmente basados en el ARN ribosomal, pero también sobre la base de proteínas (proteoma) y genes (genoma).
Históricamente se clasificaba a las bacterias desde su descubrimiento según su morfología. Posteriormente, ya en el s.XX se incluyó los aspectos bioquímicos y metabólicos. Para los años 1990 ya se disponía de catálogos nucleotídicos de ARNr 16S para más de 450 bacterias, por lo que la actual taxonomía bacteriana está muy influenciada por los estudios del ARNr.
A pesar de que existe una amplia base de datos sobre la filogenia molecular procariota, los resultados se integran al árbol de la vida solo parcialmente y se habla incluso del fracaso de tales esfuerzos. El principal factor que dificulta conocer la evolución y filogenia bacteriana es la transferencia genética horizontal, por lo que un árbol filogenético determinado dependerá del gen o genes escogidos, produciéndose resultados muy diversos, contradictorios y difíciles de interpretar. Otro factor que puede incurrir es la atracción de ramas largas, un artefacto filogenético que coloca erróneamente a los linajes de rápida evolución en posición basal o con otros linajes de rápida evolución altamente divergentes. Aun así, algunos autores han ensayado teorías evolutivas que intentan explicar la naturaleza del ancestro común bacteriano y su evolución en grandes grupos filogenéticos.
Las bacterias termófilas e hipertermófilas suelen tener algunas características comunes como la respiración anaerobia, la reducción de sulfatos o sulfuros y la mayoría son bacterias Gram negativas por su doble membrana. Pueden tener un metabolismo quimiolitoautótrofo al igual que muchas arqueas o ser quimioorganótrofos. Comúnmente se desarrollan bien entre los 60 y 90ºC.
Los árboles filogenéticos basados en el genoma, y en el proteoma muestran una clara relación entre Aquificae y Thermotogae, la cual suele extenderse a Dictyoglomi. Sin embargo el análisis ARNr 23S y 16S muestran a los termófilos como un grupo parafilético en posición basal que incluye a Thermodesulfobacteria. También se ha propuesto que la condición termófila bacteriana tendría una relación con la transferencia genética horizontal con arqueas.

Análisis sobre 56 genes extienden esta relación a Deinococcus-Thermus y análisis genómicos más actualizados agrupan a los termófilos con otros grupos recientemente descubiertos que no son necesariamente termófilos. Así pues, Aquificae se relacionaría con Chrysiogenetes, Deferribacteres y Thermodesulfobacteria; mientras que Thermotogae se aproxima más a Dictyoglomi, Caldiserica y Synergistetes. Por otro lado, análisis del proteoma agrupa a los termófilos con Nitrospirae.
Otros filos que son predominantemente mesófilos poseen subgrupos termófilos, como es el caso de Firmicutes, Actinobacteria, Chloroflexi y Bacteroidetes.
Las bacterias Gram positivas o Posibacteria, están conformadas por los filos: Firmicutes (GC bajo) y Actinobacteria (GC alto). Otro filo relacionado es Chloroflexi, por su proximidad filogenética y su estructura monérmica, aunque su tinción Gram es variable. Se considera que el filo Tenericutes (micoplasmas) son derivados de Firmicutes según la gran mayoría de árboles filogenéticos, pero su tinción no es Gram positiva debido a la carencia de pared celular.
Los Gram positivos tienen características comunes, especialmente en la envoltura celular, la cual está conformada por una sola membrana a diferencia de las dos membranas que poseen las bacterias Gram negativas. Están revestidas externamente por una gruesa pared de peptidoglicano. Son bacterias anaerobias o aerobias cuyo hábitat es principalmente continental, encontrándose tanto en el suelo como en agua dulce y muchas veces colonizando animales y plantas como comensales o como invasores patógenos. Suelen desarrollar esporas, ya que unos firmicutes producen endosporas y algunas actinobacterias producen exosporas.
La posición de Firmicutes y Actinobacteria en los árboles filogenéticos, muestra que su relación no es determinante. Forman un grupo monofilético en algunos árboles proteicos, como en el basado en la proteína EF-1α-Tu y en las proteínas de choque térmico Hsp70 y Hsp60. En otros árboles proteicos la relación evolutiva es próxima, en donde Firmicutes aparece primero, dando origen a Actinobacteria posteriormente. Por el contrario, en algunos árboles de ARNr 16S, es Actinobacteria quien aparece como un grupo anterior al clado Firmicutes. Por otro lado hay árboles genómicos que los muestran como grupos separados, pero de algún modo relacionados dentro de un clado mayor denominado Terrabacteria, por lo que sus características comunes como la gruesa pared y la formación de esporas, podrían ser producto de la adaptación al hábitat terrestre.
Los grupos fotosintéticos presentan algunas características comunes como la tinción Gram negativa, la presencia de un pigmento verde fotosintético tipo porfirina para el metabolismo fotolitoautótrofo y presencia de otros pigmentos como los carotenoides. Se diferencian principalmente en el tipo de fotosíntesis, que en Cyanobacteria por ejemplo es oxigénica (con producción de oxígeno) por acción de la clorofila, mientras que la de Chloroflexia y Chlorobia es anoxigénica por acción de la bacterioclorofila.
Se ha encontrado que Cyanobacteria y Chloroflexi tienen unidad monofilética en varios árboles genómicos y proteicos. En otros estudios genómicos y proteicos también se encuentran relacionados entre sí, pero se incluyen en un clado conjuntamente con Deinococcus-Thermus. Por el contrario, en los árboles de ARNr 16S y 23S no se ha presentado ninguna relación cercana entre estos dos grupos.
Es posible que la bacterioclorofila apareciera primero, el pasado arcaico de una atmósfera sin oxígeno parece corresponder con la fotosíntesis anoxigénica. El estudio filogenético de los diversos pigmentos concluye por consenso que la bacterioclorofila precedió a la clorofila, además la molécula de la bacterioclorofila es más simple y a la vez más diversa. El análisis de secuencias de nucleótidos que codifican las enzimas biosintéticas de la clorofila y la bacterioclorofila refuerzan este punto.
Otros grupos fotótrofos como las bacterias verdes sulfurosas, bacterias púrpuras, heliobacterias y otras, poseen bacterioclorofila, pero no están relacionados filogenéticamente con Cyanobacteria ni con Chloroflexi, por lo que probablemente adquirieron su condición fotótrofa en forma secundaria mediante transferencia genética horizontal.
En general podemos notar de acuerdo a los análisis filogenéticos, que los grupos fotosintéticos no serían los más antiguos, ya que ellos aparecen más bien como subgrupos derivados de ancestros heterótrofos: Cyanobacteria se derivaría del clado Cyanobacteria-Melainabacteria, Chloroflexia de derivaría de otros Chloroflexi, Chlorobia de otros Chlorobi, las bacterias púrpuras de otras proteobacterias, las heliobacterias de Firmicutes, etc.
Gracilicutes o Hydrobacteria, es un extenso grupo de bacterias Gram negativas, y como tales contienen doble membrana celular: Su metabolismo es quimioorganoheterótrofo en la mayoría, pues muchos dependen de materia orgánica para su subsistencia; su respiración es anaerobia o aerobia, suelen presentar flagelos, y es común la relación con animales y plantas como comensales o patógenos. Es un supergrupo constituido a su vez por cuatro grupos principales:
Gracilicutes está respaldado por la concatenación de los árboles de ARNr 16S y 23, así como en varios estudios proteicos. Igualmente hay respaldo en un estudio genómico donde se analiza la inserción y eliminación de secuencias proteicas, relacionando además a Aquificae con Proteobacteria. Otro estudio genómico apoya ampliamente este clado, pero contradiciendo otros estudios ubica a Aquificae junto Thermotogae y fuera de Gracilicutes, ya que se encontró evidencia de una gran transferencia genética horizontal entre los aquificales y épsilon-Proteobacteria. Un extenso análisis del ARNr 16S respalda este clado e incluye a Fusobacteria y a pequeños filos recién descubiertos como Chrysiogenetes y Armatimonadetes.

Evolutivamente, Gracilicutes se habría originado a partir de una bacteria Gram negativa flagelada poseedora de lipopolisacáridos (endotoxinas) en sus membranas, la cual sufrió cuatro inserciones proteicas: un amino-acil en Hsp60 y en FtsZ, y un dominio en alfa y en beta ARNP.
Es probable que la primera bacteria haya heredado características hipertermófilas, quimiótrofas y anaerobias del último ancestro universal, pues esto es compatible con las hipótesis más aceptadas sobre el origen de la vida: la del "caldo primigenio" y la teoría del mundo de hierro-azufre.
Los árboles filogenéticos basados en el ARNr 16S y 23S, muestran a las bacterias termófilas en posición basal; al igual que en algunos árboles proteicos y enzimáticos. En la gran mayoría de estos casos, el filo Aquificae es la rama bacteriana más antigua y divergente, un filo cuyo metabolismo quimiosintético recuerda a muchas arqueas. En cuanto a los árboles genómicos, en su mayoría dan otros resultados y no avalan estas conclusiones.
La concatenación de los árboles filogenéticos de ARNr 16S y 23S ha dado el siguiente resultado:
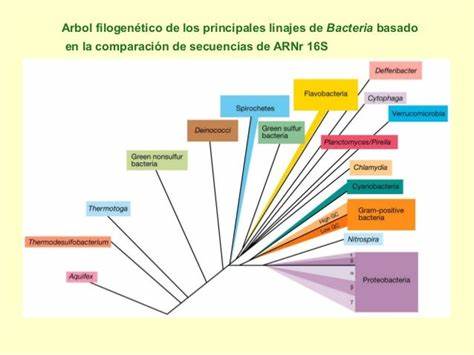

Otros análisis en base ARNr 16S y de 23S, coinciden en dar como grupos basales a Aquificae (un grupo hipertermófilo quimiosintético) y Thermotogae (termófilo quimioheterótrofo).
Un extenso análisis basado en el ARNr 16S (proyecto The All-Species Living Tree), con una actualización en la filogenia profunda (2018), da el siguiente resultado (los grupos parafiléticos van entre comillas):
"Firmicutes" (incluye Cyanobacteria, Tenericutes y Dictyoglomi)
Chloroflexi (inc. Thermomicrobia)
Este árbol da como resultado al menos 5 grupos termófilos-quimiótrofos-anaerobios bacterianos en posición basal. Si a esto sumamos que la mayoría de filos de las arqueas son igualmente termófilos-quimiótrofos-anaerobios, podemos apreciar que la filogenia procariota nos ha dado así, una idea de las características de la vida durante los albores de la historia de la Tierra. Adicionalmente se observa que los filos basales Coprothermobacterota y Thermotogae se consideran monodérmicos, con una sola membrana lipídica como las arqueas.
Más ancestros termófilos encontramos al observar la filogenia de otros filos según este mismo proyecto del ARNr 16S. Los subgrupos basales termófilos son: Thermaerobacter, Thermoanaerobacteraceae, Dictyoglomi y Sulfobacillus (subclados de Firmicutes), Rubrobacteridae (Actinobacteria) y Rhodothermaceae (Bacteroidetes). También son termófilos algunos subgrupos de Chloroflexi, Deinococcus-Thermus, Nitrospirae, Chlorobi, Deferribacteres, Acidobacteria, el género Caldithrix y algunas especies de las protebacterias delta y beta.
Algunos estudios filogenéticos proteicos coinciden en que los grupos termófilos son los más divergentes. Además muestran que entre las demás bacterias (las no termófilas) hay dos grandes supergrupos, uno adaptado al hábitat terrestre denominado Terrabacteria y otro adaptado al hábitat marino llamado Hydrobacteria (bacteria de agua) o Gracilicutes (de piel grácil por su delgada pared). Terrabacteria muestra varias características relacionadas con la adaptación al hábitat terrestre, como la resistencia a la desecación, la formación de esporas y la aparición de pigmentos que proveen protección de la radiación solar, lo que pudo ocurrir desde hace unos 3.000 millones de años.
Terrabacteria se descubrió sobre la base de un estudio sobre 32 proteínas en 72 especies que mostró que había un superclado que agrupaba a Firmicutes, Actinobacteria, Cyanobacteria, Deinococcus-Thermus y Fusobacteria (Battistuzzi 2004). Estudios proteicos posteriores con similares resultados, relacionan a Cyanobacteria con Chloroflexi, y estos a su vez con los grupos Gram positivos. Estos resultados han llevado a conclusiones evolutivas: las bacterias Gram positivas se habrían originado a través de la adaptación al hábitat terrestre, con una gruesa pared que las protege de la desecación y que ha contribuido a la patogenicidad en muchas especies. Por lo general, los árboles evolutivos procariotas se basan en filos y clases, considerando que las relaciones entre estos grupos son producto de la transferencia horizontal de genes, sesgos en el cambio de secuencias y a grandes distancias evolutivas; pero rara vez se apoyan en la ecología y la biología celular. El siguiente "anillo de la vida", es en realidad un árbol ecológico simplificado que combina los resultados obtenidos por estos y otros autores, basándose en la evidencia filogenética, citológica y medioambiental:
Algunos análisis genómicos también colocan a los grupos hipertermófilos en posición basal. En forma resumida se muestra el siguiente cladograma:
supergrupo Terrabacteria
supergrupo Gracilicutes o Hydrobacteria
El clado Selabacteria antecede a la aparición de los primeros grupos fotótrofos hace más de 3000 millones de años.
El supergrupo CPR (Candidate Phyla Radiation) de las bacterias ultrapequeñas o nanobacterias, descubierto en 1998 y descrito recientemente en 2015, es un nuevo tipo de vida enigmático que se encontraría en la base de la filogenia bacteriana de acuerdo con nuevos métodos de análisis que estudian las proteínas ribosómicas. Las bacterias CPR son generalmente simbiontes de pequeño genoma con capacidades metabólicas que van de algo limitadas a muy restringidas, sin ciclo del ácido cítrico ni cadenas respiratorias y con limitada o ninguna capacidad para sintetizar nucleótidos y aminoácidos. El análisis genómico basado en proteínas ribosómicas ha dado la siguiente filogenia:
Cyanobacteria / Melainabacteria
Firmicutes (inc. Tenericutes, Armatimonadetes)
Acidobacteria y relacionados
Thermodesulfobacteria y relacionados
Chrysiogenetes / Deferribacteres
Hydrogenedentes / Spirochaetes
Si se compara el análisis fitogenético de las proteínas ribosómicas con otros estudios moleculares (ARNr, proteínas, genes), así como con teorías evolutivas, se obtiene los siguientes resultados aproximados:
Un estudio genómico que compara múltiples árboles proteicos y genes homólogos (2001), descubre tres potenciales clados bacterianos del más alto nivel filogenético:
Estos tres grupos coinciden con los supergrupos Gracilicutes, Terrabacteria y el de los hipertermófilos respectivamente.
También es posible que en Bacteria se produjo una temprana divergencia en dos grandes superclados: Esta hipótesis está respaldada por el análisis filogenético de un grupo representativo de proteínas. El resultado del análisis de 56 genes bacterianos respaldan este resultado, llamando a estos clados Terrabacteria y Gracilicutes, obtiendo las siguientes relaciones (2015):
Un análisis genómico (2020) usando genes como caracteres y teniendo en cuenta la transferencia horizontal y la tasa de mutación también apoya este modelo incluyendo a las bacterias ultrapequeñas (CPR) en Terrabacteria, mientras que el filo Fusobacteria puede pertenecer alternativamente a Terrabacteria, Gracilicutes o incluso puede ser el filo más basal de todas las bacterias. Se ha obtenido aproximadamente el siguiente resultado.
DST (Deinococcus-Thermus, Synergistetes, Thermotogae y relacionados)
Fusobacteria (?)
FCB (Fibrobacteres, Cloacimonetes, Bacteroidetes y relacionados)
PVC (Planctomycetes, Verrucomicrobia, Chlamydiae y relacionados)
Según R. Gupta, es posible que el último ancestro universal fuese una bacteria Gram positiva debido a su estructura simple y monodérmica, que al igual que las arqueas presentan una sola membrana celular. Las bacterias gramnegativas se habrían originado a partir de una grampositiva que adquirió la membrana externa como protección contra el medio externo, en especial contra antibióticos producidos por otras Gram positivas.
Algunos árboles genómicos como el de la derecha, presentan a Firmicutes como un clado basal. El árbol filogenético de abajo, está basado en genes codificadores de proteínas (2005).
De acuerdo con Gupta, [A] (mostrado en este cladograma), representa evolutivamente la adquisición de la membrana externa; [B] es la inserción de la proteína Hsp70, y [C] representa el clado de bacterias didérmicas con inserción de la proteína Hsp60 y la evolución de los genes de la biosínteses de lipopolisacáridos (LPS).
Bajo este punto de vista, habría dos tipos de bacterias didérmicas gramnegativas: las didérmicas típicas con Hsp60 y LPS representadas por los clados Gracilicutes y Cyanobacteria/Melainabacteria; y las didérmicas atípicas representadas por los filos Thermotogae, Fusobacteria, Synergistetes, Elusimicrobia, Deinococcus-Thermus y otros.
El descubrimiento de las bacterias ultrapequeñas, las cuales son consideradas primitivas y mayormente monodérmicas, podría significar un respaldo a la hipótesis del ancestro monodérmico.
La siguiente figura muestra un árbol filogenético de los seres vivos basado en las ideas de Cavalier-Smith. Según este autor, la raíz del árbol se situaría entre las bacterias Gram-negativas, pues los organismos más antiguos serían fotosintéticos y similares a las bacterias verdes no del azufre (existiendo desde hace 3.500 millones de años), mientras que Archaea y Eukaryota serían relativamente recientes (de hace solo 900 millones años).
El árbol se basa en la estructura celular de los distintos seres vivos enfatizando en la envoltura celular (membrana citoplasmática, pared celular y membrana externa). Según este criterio, el dominio Bacteria contiene organismos con dos tipos distintos de organización básica, Gram-negativa y Gram-positiva, y además podemos subdividir a las Gram-negativas en dos subgrupos en función de la composición de la membrana externa.
Negibacteria (bacterias Gram negativas) presenta dos membranas lipídicas distintas, entre las que se localiza la pared celular, mientras que el resto de los organismos presentan una única membrana lipídica. La hipótesis del citoplasma fuera describe un posible modelo para la aparición de las dos membranas en estas primeras bacterias. Dentro de este grupo podemos distinguir dos subgrupos. Los subgrupos Eobacteria y Glycobacteria se distinguen por la composición de la membrana externa, que presenta solo simples fosfolípidos en los primeros e inserción de moléculas complejas de lipopolisacáridos en los segundos.
Posibacteria (bacterias Gram positivas) presenta una única membrana y la pared de peptidoglicano (mureína) se hace mucho más gruesa. Se considera que las posibacterias proceden de las negibacterias, y no al revés, porque las primeras presentan características moleculares y ultraestructurales más avanzadas. La pérdida de la membrana externa podría ser debida a la hipertrofia de la pared celular, que aumenta la resistencia de estos organismos, pero impide la transferencia de lípidos para formar la membrana externa. Estos organismos fueron probablemente los primeros que colonizaron el suelo.
Archaea y Eukarya probablemente tuvieron como origen una Posibacteria a través de un organismo Neomura que sustituyó la pared celular de peptidoglucano por otra de glicoproteína. A continuación y casi inmediatamente, las arqueas se adaptaron a ambientes calientes y ácidos, reemplazando los lípidos acilo éster de las bacterias por lípidos prenil éter, y usaron las glicoproteínas como una nueva pared rígida. Los eucariontes, en cambio, usaron la nueva superficie de proteínas como una capa flexible para desarrollar la fagocitosis, lo que los llevó, en última instancia, a profundos cambios en la estructura de la célula.
Cavalier-Smith no reconoce a Archaea como un dominio ya que este descendería de bacterias, por lo que usa el término anterior Archaebacteria. El siguiente cladograma muestra más en detalle las relaciones entre los distintos grupos de seres vivos en donde las bacterias tienen un papel central, de acuerdo con las ideas de Cavalier-Smith:
Leyendas:
Cavalier-Smith (2020) en un análisis de múltiples proteínas ribosómicas ha encontrado soporte para su hipótesis con Cloroflexi como el clado más basal de Cytota y las bacterias siendo parafiléticas de Neomura, pero este árbol difería del anterior en que el ancestro de Neomura estaría entre Planctobacteria, Sphingobacteria o Thermobacteria y las arqueas eran parafiléticas de los eucariotas como lo sugirieron los otros análisis moleculares. Por lo que ahora una versión revisada reconoce el origen de los eucariotas dentro de Archaea y propone que el ancestro más probable de este grupo sea una planctobacteria y no una actinobacteria como se propuso anteriormente. Este análisis podría ser un respaldo al ancestro fotosintético y también sugiere que las bacterias como dominio pueden ser directamente un taxón parafilético.
El árbol actualizado de Cavalier-Smith es aproximadamente el siguiente:
Sphingobacteria (FCB)
Planctobacteria (PVC)
Euryarchaeota (incluye a DPANN)
Crenarchaeota s.l (TACK)
Diversos estudios dan resultados muy diferentes. Muchos de ellos no reconocen a Gracilicutes como un supergrupo y son diversos los filos que pueden encontrarse en posición basal. Por ejemplo, en la filogenia de la proteína estructural FtsZ, aparece en posición basal Alphaproteobacteria. Un estudio sobre secuencias del ARNr sostiene que es Planctomycetes el filo más divergente, aun cuando lo usual en los árboles ARNr es la temprana divergencia de los termófilos. Igualmente un estudio del proteoma coloca a Planctomycetes en esta posición. En otros estudios encontramos en posición basal a Deinococcus-Thermus según genes codificadores de proteínas, Spirochaetes según 730 genes, Proteobacteria según marcadores genéticos, Cyanobacteria o Bacteroidetes según secuencias de ARNP, Chlamydiae según la proteína Hsp60 y Tenericutes según la enzima aminoacil ARNt sintetasa.
Todos estos árboles presentados, separan claramente a Bacteria de los otros dominios Archaea y Eukaryota. Sin embargo, otros árboles como los de las enzimas ATPasa o girasa no existe una organización en tres dominios.
Muchos de estos grupos que aparecen como los más divergentes, serían producto de cambios drásticos en el genoma. Por ejemplo, en Tenericutes, Alphaproteobacteria, Chlamydiae y CPR ha habido una importante reducción y/o cambios en el genoma ocasionados por el modo de vida parasitario-endosimbiótico; o cambios relacionados con la complejidad estructural en Spirochaetes y Planctomycetes. Así pues, sugerir que estos grupos son los más antiguos, podría significar caer en un error por el factor de atracción de ramas largas.
Escribe un comentario o lo que quieras sobre Filogenia bacteriana (directo, no tienes que registrarte)
Comentarios
(de más nuevos a más antiguos)
Aún no hay comentarios, ¡deja el primero!